



| Biochemistry has confirmed and greatly
extended evolutionary theory. Carolus Linnaeus recognized
the anatomic similarities and differences among living
organisms and provided a framework for assessing the
relatedness of different species. Charles Darwin gave us
a unifying hypothesis to explain the phylogeny of modern
organisms-the origin of different species from a common
ancestor. Biochemistry has begun to reveal the molecular
anatomy of cells of different species-the sequences of
subunits in nucleic acids and proteins and the
three-dimensional structures of individual molecules of
nucleic acid and protein. There is a reasonable prospect
that when the twenty-first century dawns, we will know
the entire nucleotide sequence of all of the genes that
make up the biological heritage of a human. At the molecular level, evolution is the emergence over time of different sequences of nucleotides within genes. With new genetic sequences being experimentally determined almost daily, biochemists have an enormously rich treasury of evidence with which to analyze evolutionary relationships and to refine evolutionary theory. The molecular phylogeny derived from gene sequences is consistent with, but in many cases more precise than, the classical phylogeny based on macroscopic structures. Molecular structures and mechanisms have been conserved in evolution even though organisms have continuously diverged at the level of gross anatomy. At the molecular level, the basic unity of life is readily apparent; crucial molecular structures and mechanisms are remarkably similar from the simplest to the most complex organisms. Biochemistry makes it possible to discover the unifying features common to all life. This book examines many of these features: the mechanisms for energy conservation, biosynthesis, gene replication, and gene expression. |

|
| The information in DNA is encoded as a
linear (one-dimensional) sequence of the nucleotide units
of DNA, but the expression of this information results in
a three-dimensional cell. This change from one to three
dimensions occurs in two phases. A linear sequence of
deoxyribonucleotides in DNA codes (through the
intermediary, RNA) for the production of a protein with a
corresponding linear sequence of amino acids (Fig. 1-18).
The protein folds itself into a particular
three-dimensional shape, dictated by its amino acid
sequence. The precise threedimensional structure (native
conformation) is crucial to the protein's function as
either catalyst or structural element. This principle
emerges : The linear sequence of amino acids in a protein leads to the acquisition of a unique three-dimensional structure by a self assembly process. Once a protein has folded into its native conformation, it may associate noncovalently with other proteins, or with nucleic acids or lipids, to form supramolecular complexes such as chromosomes, ribosomes, and membranes (Fig. 1-18). These complexes are in many cases self assembling. The individual molecules of these complexes have specific, high-affinity binding sites for each other, and within the cell they spontaneously form functional complexes. Individual macromolecules with specific affinity for other macromolecules self assemble into supramolecular complexes. |
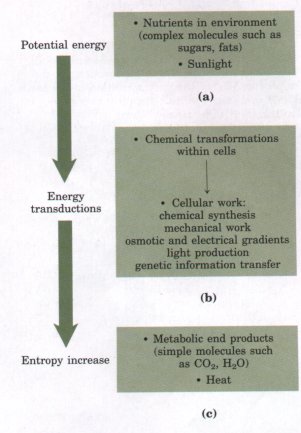 Fignre 1-18 Linear sequences of deoxyribonucleotides in DNA, arranged into units known as genes, are transcribed into ribonucleic acid (RNA) molecules with complementary ribonucleotide sequences. The RNA sequences are then translated into linear protein chains, which fold spontaneously into their native three-dimensional shapes. Individual proteins sometimes associate with other proteins to form supramolecular complexes, stabilized by numerous weak interactions. |
The forces that provide stability and specificity to the three-dimensional structures of macromolecules and supramolecular complexes are mostly noncovalent interactions. These interactions, individually weak but collectively strong, include hydrogen bonds, ionic interactions among charged groups, van der Waals interactions, and hydrophobic interactions among nonpolar groups. These weak interactions are transient; individually they form and break in small fractions of a second. The transient nature of noncovalent interactions confers a flexibility on macromolecules that is critical to their function. Furthermore, the large number of noncovalent interactions in a single macromolecule makes it unlikely that at any given moment all the interactions will be broken; thus macromolecular structures are stable over time.
Three-dimensional biological structures combine the properties of flexibility and stability.
The flexibility and stability of the double-helical structure of DNA are due to the complementarity of its two strands and the many weak interactions between them. The flexibility of these interactions allows strand separation during DNA replication (see Fig. 1-16); the complementarity of the double helix is essential to genetic continuity.
Noncovalent interactions are also central to the specificity and catalytic efficiency of enzymes. Enzymes bind transition-state intermediates through numerous weak but precisely oriented interactions. Because the weak interactions are flexible, the complex survives the structural distortions as the reactant is converted into product.
The formation of noncovalent interactions provides the energy for self assembly of macromolecules by stabilizing native conformations relative to unfolded, random forms. The native conformation of a protein is that in which the energetic advantages of forming weak interactions counterbalance the tendency of the protein chain to assume random forms. Given a specific linear sequence of amino acids and a specific set of conditions (temperature, ionic conditions, pH), a protein will assume its native conformation spontaneously, without a template or scaffold to direct the folding.
We can now summarize the various principles of the molecular logic of life:
A living cell is a self contained, self assembling, self adjusting, self perpetuating isothermal system of molecules that extracts free energy and raw materials from its environment.
The cell carries out many consecutive reactions promoted by specific catalysts, called enzymes, which it produces itself.
The cell maintains itself in a dynamic steady state, far from equilibrium with its surroundings. There is great economy of parts and processes, achieved by regulation of the catalytic activity of key enzymes.
Self replication through many generations is ensured by the self repairing, linear information-coding system. Genetic information encoded as sequences of nucleotide subunits in DNA and RNA specifies the sequence of amino acids in each distinct protein, which ultimately determines the three-dimensional structure and function of each protein.
Many weak (noncovalent) interactions, acting cooperatively, stabilize the three-dimensional structures of biomolecules and supramolecular complexes.
At no point in our examination of the molecular logic of living cells have we encountered any violation of known physical laws; nor have we needed to define new physical laws. The organic machinery of living cells functions within the same set of laws that governs the operation of inanimate machines, but the chemical reactions and regulatory processes of cells have been highly refined during evolution.
This set of principles has been most thoroughly validated in studies of unicellular organisms (such as the bacterium E. coli), which are exceptionally amenable to biochemical and genetic study. Although multicellular organisms must solve certain problems not encountered by unicellular organisms, such as the differentiation of the fertilized egg into specialized cell types, the same principles have been found to apply. Can such simple and mechanical statements apply to humans as well, with their extraordinary capacity for thought, language, and creativity? The pace of recent biochemical progress toward understanding such processes as gene regulation, cellular differentiation, communication among cells, and neural function has been extraordinarily fast, and is accelerating. The success of biochemical methods in solving and redefining these problems justifies the hope that the most complex functions of the most highly developed organism will eventually be explicable in molecular terms.
The relevant facts of biochemistry are many; the student approaching this subject for the first time may occasionally feel overwhelmed. Perhaps the most encouraging development in twentieth century biology is the realization that, for all of the enormous diversity in the biological world, there is a fundamental unity and simplicity to life. The organizing principles, the biochemical unity, and the evolutionary perspective of diversity, provided at the molecular level, will serve as helpful frames of reference for the study of biochemistry.
Asimov, I. (1962) Life and Energy: An Exploration of the Physical and Chemical Basis of Modern Biology, Doubleday & Co., Inc., New York.
An engaging account of the role of energy transformations in biology, written for the intelligent layman.
Blum, H.F. (1968) Time's Arrow and Euolution, 3rd edn, Princeton University Press, Princeton, NJ.
An excellent discussion of the way the second law of thermodynamics has influenced biological euolution.
Dulbecco, R. (1987) The Design of Life, Yale University Press, New Haven, CT.
An unusual and excellent introduction to biology.
Fruton, J.S. (1972) Molecules and Life. Historical Essays on the Interplay of Chemistry and Biology, Wiley-Interscience, New York.
This series of essays describes the deuelopment of biochemistry from Pasteur's studies of fermentation to the present studies of metabolism and information transfer. You may want to refer to these essays through this textbook.
Fruton, J.S. (1992) A Skeptical Biochemist, Harvard University Press, Cambridge, MA.
Hawking, S. (1988) A Brief History of Time, Bantam Books, Inc., New York.
Jacob, F. (1973) The Logic of Life: A History of Heredity, Pantheon Books, Inc., New York. Originally published (1970) as La logique du vivant: une histoire de L'heredite, Editions Gallimard, Paris.
A fascinating historical and philosophical account of the route by which we came to the present molecular understanding of life.
Kornberg, A. (1987) The two cultures: chemistry and biology. Biochem. 26, 6888-6891.
The importance of applying chemical tools to biological problems, described by an eminent practitioner.
Monod, J. (1971) Chance and Necessity, Alfred A. Knopf, Inc., New York. [Paperback version (1972) Vintage Books, New York.] Originally published (1970) as Le hasard et la necessite, Editions du Seuil, Paris.
An exploration of the philosophical implications of biological knowledge.
Schrodinger, E. (1944) What is Life? Cambridge University Press, New York. [Reprinted (1956) in What is Life? and Other Scientific Essays, Doubleday Anchor Books, Garden City, NY.]
A thought-provoking look at life, written by a prominent physical chemist.

